Artikel - Genförluster hos sällskapsdjur - ett hälsoproblem?
GENFÖRLUSTER
HOS SÄLLSKAPSDJUR - ETT HÄLSOPROBLEM?
Per-Erik Sundgren
Inst. för husdjurgenetik
SLU
Sammanfattning
• Villkoren
för avel med sällskapsdjur har förändrats dramatiskt de senaste 100 åren.
Hund- och katt- utställningar har skapat en tävlingsverksamhet som i många
fall drivit fram intensiv avel med extrema avelsmål i mycket små djurpopulationer.
• Genomförda analyser visar att den effektiva populationens storlek (ett
mått på takten i genförluster) i de flesta raser motsvarar mindre än 100 djur
i slumpparning, trots att det faktiska djurantalet i några av dem uppgår till
tiotusentals individer
• Studier av DNA-sekvenser, sk mikrosatelliter, och plasmaproteiner, i 6 utvalda hundraser visar starka samband mellan beräknad inavelsgrad och förlust av ärftlig variation.
• De
raser i studien som har den lägsta ärftliga variationen i DNA och genprodukter
är också kända för relativt
omfattande ärftliga problem. Rasernas faktiska numerär visar stor skillnad
men den effektiva populationsstorleken är i båda fallen liten.
• En kombination av DNA-analys och en skattning av den effektiva populationsstorleken
kan vara en väg att snabbt kartlägga vilka raser som befinner sig i den omedelbara
riskzonen för allvarliga genetiska förluster.
• Det är nödvändigt med ett nytänkande kring tävlingsformer och avelsplanering i ett stort antal raser av sällskapsdjur om de på sikt skall kunna hållas rimligt friska och fria från ärftliga störningar.
Bakgrund
Det är känt att många raser av sällskapsdjur har bildats med utgångspunkt
från ett litet antal individer. Av tradition har hundaveln i stor utsträckning
bedrivits som linjeavel, ofta med omfattande utnyttjande av nära släktskapsavel.
Koncentrationen av avelsarbete kring få individer har förstärkts av de premieringssystem
som tillämpas i avel med sällskapsdjur, främst inom utställningsverksamheten
med dess konkurrensbedömningar.
I flera länder har den ökande frekvenser av genetiska skador hos både lantbrukets husdjur och sport- och fritidsdjur uppmärksammats. Lagstiftarna har reagerat genom att införa lagbestämmelser avsedda att begränsa sådana former av avelsarbete som kan skapa djurlidanden. Kunskapen om vilka åtgärder som krävs för att undvika genetiska skador som en följd av felaktig avelsplanering finns främst inom den traditionella forskningen kring lantbrukets husdjur. Problem som en följd av avel i små populationer kommer ändå, med utnyttjande av modern fortplantningsteknik, att i ökande utsträckning göra sig gällande också vid avel med lantbrukets husdjur. Forskning kring hund kan bidra med intressant kunskap inom området. Den fortsatta redovisningen gäller främst studier på hund.
De forskningsresultat som här redovisas från DNA- och plasmaproteinstudier från sex hundraser har tagits fram i samverkan främst med Norsk Kennelklubbs forskningschef Frode Lingaas, verksam vid Norges Veterinärhögskola, och agr.dr. Kumar Juneja, verksam vid Institutionen för husdjursförädling och sjukdomsgenetik, SLU. Studierna har möjliggjorts tack vare finansiellt stöd från Svenska Kennelklubben, försäkringsbolaget Agria och Finska Kennelklubben samt genom medverkan från ett stort antal veterinärer och hundägare i Sverige.
Raser i
slutna populationer är en sen företeelse
Många hundraser har en mycket lång utvecklingsbakgrund. Det finns historiska
belägg för att hundstammar, med i stora drag likartat utseende som dagens
hundar, funnits under århundraden eller rentav årtusenden. Förbättrade kommunikationer
och introduktionen av rasbegreppet i dess moderna form, och den tävlingsverksamhet
detta lett fram till, har emellertid åstadkommit dramatiska förändringar i
avelsarbetets villkor. Med förbättrade kommunikationer kan en och samma hanhund
utnyttjas för parning med tikar över ett långt mycket större område än tidigare.
Färre hanhundar behövs därmed för rasens fortplantning och var och en kan
få ett avsevärt större antal avkomlingar.
Föreställningar
om rasrenhetens betydelse för individers kvalitet förstärktes betydligt under
det sena 1800-talet och den första tredjedelen av 1900-talet. På den grunden
skapades starka restriktioner i avelsarbetet med strikt slutna stamböcker
för populationer som ofta grundats på bara ett fåtal individer. Den på rasbegreppet
grundade tävlingsverksamheten vid utställningar, som tog sin egentliga början
i slutet av 1800-talet, skapade grunden för ett mycket intensivt urval för
bestämda exteriöra typer med åtföljande stark koncentration av aveln kring
ett fåtal individer. Som ett resultat av de beskrivna förändringarna i avelsarbetets
villkor har inavelsökningen i många stammar av sällskapsdjur under 1900-talet
fortgått i en takt som i tidigare skeden aldrig var vare sig möjlig eller
aktuell.
Effektiva och skenbara populationer
En ras utgörs av det antal djur som av en organisation, exempelvis en kennelklubb, bokförs under en gemensam beteckning och där villkoret som regel är att varje djur som bokförs skall ha föräldrar bokförda under samma rasbeteckning. Rasen bildar därmed en sluten grupp djur inom vilken all avel bedrivs. Utöver detta definieras sedan rasen genom att varje ras ges en relativt ingående exteriör beskrivning. För jakt- och brukshundraser finns dessutom normalt en funktionsbeskrivning sammanfattad i jakt- eller bruksprovsregler. Alla individer som bokförts som rasrena, dvs är fallna efter bokförda föräldrar, är potentiella avelsdjur. De faktiska avelsdjuren väljs dock i stor utsträckning ur den grupp djur som deltagit i tävlingar och prov och därvid beskrivits som rastypiska enligt gällande exteriörstandard eller vunnit vid konkurrensbedömningar. Detta leder till att långtifrån hela djurpopulationen utnyttjas i avel. Därmed uppstår en skillnad mellan det faktiska antalet djur som finns i rasen och den egentliga avelspopulationens storlek.
Inom avelsläran används ett begrepp, effektiv populationsstorlek , för att tydliggöra skillnaden mellan tillgången till individer inom en djurstam och det sätt på vilket individerna utnyttjas i avelsarbetet. Som en följd av inavel eller slumpens krafter, sk genetic drift , förloras genmaterial olika snabbt i populationer av olika storlek. Ett sätt att beskriva takten i genförlusterna är att beräkna inavelsgradens stegring per generation, ?F, i en djurstam. Förändringen, ?F, anger då hur stor del av den kvarvarande ärftliga variationen i stammen som går förlorad på en generation. Sådana beräkningar kan göras i verkliga populationer med ledning av härstamningsuppgifter ur respektive stamböcker.
Beräkningar av ?F utförda på verkliga populationer kan sedan jämföras mot genförluster i idealiserade populationer. En idealiserad population är en population där all parning är slumpmässig och där inget urval sker. Mutationen påverkar inte genfrekvenserna. Formellt betraktas dessutom alla individer som självbefruktande. Vid jämförelser på husdjur är detta inte relevant för en enstaka generation, men den genetiska effekten över flera generationer blir densamma som för en idealiserad population. I en idealiserad population gäller att förlusten av ärftlig variation som en följd av rena slumpkrafter är omvänt proportionell mot antalet individer i populationen, dvs ?F= 1/2N. Detta kan också uttryckas så att N= 1/(2 ?F). Genom att beräkna ?F i verkliga populationer kan man därmed skatta hur stor den idealiserade population måste vara där genförlusten sker i samma takt som i den verkliga populationen. Den beräknade storleken kallas då den effektiva populationsstorleken och betecknas Ne och definieras som Ne= 1/(2 ?F). Detta kan också i ord
uttryckas så att den effektiva populationen motsvarar det antal djur som, under slumpparning och vid lika könsfördelning , skulle krävas för att uppnå samma stegring av inaveln (genförlust) per generation som den verkliga populationen uppvisar.
Inom vissa
raser förekommer relativt starka grupperingar, avelslinjer eller konkurrerande
uppfödargrupper, som inte delar avelsmaterial i någon större omfattning. Inom
var och en av dessa grupper kan inavelsstegringen per generation vara relativt
hög. Även om varje grupp förlorar ärftlig variation i den takt som inavelsstegringen
för gruppen beskriver så förloras inte samma gener i var och en av grupperna.
Rasen som helhet kommer därför att förlora mindre i ärftlig variation än en
enkel beräkning av den genomsnittliga inavelsgradens ökning i stammen ger
vid handen. För att kompensera för detta har en ny typ av beräkningar genomförts.
Ett slumpvis antal kullar har valts ut i varje ras. Deras föräldradjur har
identifierats. I datorsimulering har föräldradjuren slumpvis parats med varandra
oberoende av den linjeavel eller de grupperingar som funnits i rasen. Därefter
ha inaveln beräknats för de fiktiva kullar som skapats och ett nytt ?F skattas.
Detta värde har så utnyttjats för att skatta den potentiella effektiva populationen.
Jämförelser mellan realiserad och potentiell effektiv population ger en antydan
om det finns en större potential för bevarande av gener i hundstammen än en
enkel skattning av den genomsnittliga inaveln antyder.
Studier av ärftlig variation i sex hundraser
Sex hundraser valdes för att insamla verkliga data över betydelsen av avelsprogrammens utformning och populationernas storlek på förlust av ärftlig variation och djurens hälsa. Från var och en av raserna valdes ca 30 individer, företrädesvis individer födda 1990, genom slumpsampling. För de antalsmässigt små raserna blev det nödvändigt att sampla över en något längre tidsperiod för att urvalet skulle bli rimligt representativt. För att maximera information i de relativt små sample som blev aktuella genomfördes samplingen med den restriktionen att helsyskon i möjligaste mån skulle undvikas. Omfattningen och rasval i det slutliga materialet framgår av tabell 1.
Tabell 1. Antal hundar per ras i studien och deras spridning över tiden
Ras |
Antal |
Tidsperiod |
Födda 1990 |
1. Finsk stövare |
29 |
1990 |
772 |
2. West highland white terrier |
30 |
1990-1991 |
544 |
3. Pointer |
33 |
1990-1991 |
137 |
4. Tibetansk terrier |
29 |
1990-1991 |
196 |
5. Italiensk vinthund |
27 |
1989-1991 |
43 |
6. Clumber spaniel |
32 |
1988-1991 |
42 |
Två huvudkriterier låg till grund för val av raser, antal födda valpar per år och rasernas hälsotillstånd med hänsyn till kända genetiska störningar enligt veterinär erfarenhet. Valet gjordes ur tre storleksgrupper, över 500 födda, 100-500 födda samt under 100 födda per år. Raserna 1, 3 och 5 enligt tabell 1 var kända för gott hälsotillstånd. Raserna 2, 4 och 6 ansågs vid urvalstillfället mer än genomsnittligt drabbade av ärftliga störningar såvitt var känt från veterinärmedicinska erfarenheter.
Inga samband mellan rasers verkliga och effektiva storlek
Antalet födda hundar samt realiserade och potentiella effektiva populationer enligt tidigare definitioner redovisas för de sex hundraserna i diagram 1. Där finns tre staplar per ras. En som visar antalet registrerade individer per år i rasen. Om detta antal multipliceras 7-8 ggr får man en rimlig uppfattning om det totala antalet individer i rasen. För en ras, finsk stövare är den potentiella effektiva populationen mindre än den realiserade. Det har emellertid en beräkningsteknisk bakgrund, som kommer att justeras till en slutlig rapport. För härstamningsuppgifter har utnyttjats Svenska Kennelklubbens dataregister. Inom rasen finsk stövare är importer från Finland vanliga. Finska hundar, som inte registrerats i Sverige, har därför inte kommit med i simuleringen, vilket naturligtvis sänker den effektiva populationens storlek.
Det finns ett visst samband mellan den totala populationsstorleken och de beräknade värdena för effektiva populationer, men sambandet är svagare än väntat. Den faktiska populationen är för finska stövare exempelvis ca 18 ggr större än för italiensk vinthund medan skillnaden i effektiv populationsstorlek begränsas till ca 3,5 ggr.
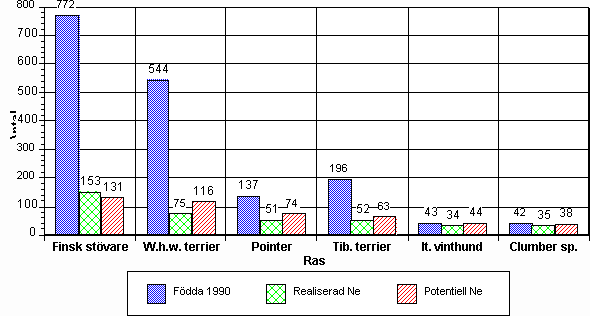
Diagram 1. Årliga registreringar samt effektiva populationer för sex hundraser
Analyser av DNA och plasmaproteiner visar god överensstämmelse
Blodprov insamlades från samtliga individer för analys av DNA och plasmaproteiner. DNA-analy-serna genomfördes vid Norges Veterinärhögskola av veterinär Frode Lingaas och proteinanalyserna vid SLU av agr.dr Kumar Juneja. Ärftlig variation, mätt som heterozygotigrad för blodanalyser, framgår av tabell 2, där också takten i inavelsgradens stegring per generation, ?F , redovisas.
Tabell 2. Total heterozygotigrad för nio plasmaproteiner och 12 mikrosatelliter samt skattad inavelsstegring per generation
Ras |
Plasmaproteiner |
Mikrosatelliter |
?F |
|
Finsk stövare |
0,30 |
0,50 |
0,33 |
|
West highland white terrier |
0,22 |
0,40 |
0,67 |
|
Pointer |
0,30 |
0,58 |
0,98 |
|
Tibetansk terrier |
0,37 |
0,61 |
0,96 |
|
Italiensk vinthund |
0,32 |
0,52 |
1,47 |
|
Clumber spaniel |
0,21 |
0,40 |
1,43 |
|
Uppmätt ärftlig variation för de studerade raserna visas i diagram 2. Diagrammet visar på en nästan total följsamhet mellan skattning av ärftlig variation direkt på DNA-nivå respektive på genprodukter. Nivåskillnaden torde främst sammanhänga med att man vid valet av mikrosatelliter för DNA-studierna valde system med ärftlig variation i samtliga raser. De studerade systemen kan därför förväntas ligga över genomsnittlig total variation för mikrosatelliter. De båda metoderna bekräftar varandra och ger hög tillförlitlighet till studiens skattning av skillnader i tillgänglig ärftlig variation mellan raser och individer.
Starka samband mellan inavel och förlust av ärftlig variation
Den totala inavelsgraden för enskilda individer beräknades över 5 generationer och sattes i relation till genomsnittlig uppmätt heterzygoti i DNA och för plasmaproteiner för varje individ. Resultatet framgår av följande digram.
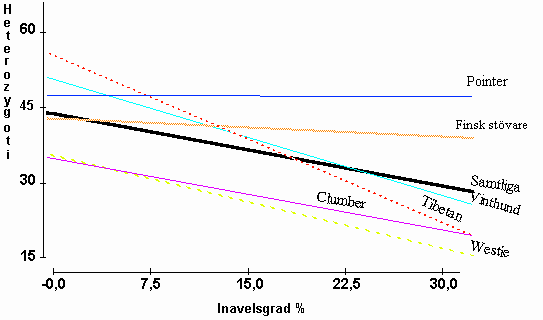
Diagram 3. Samband mellan beräknade grader av inavel och heterozygoti för enskilda hundar inom sex hundraser.
Regressionskoefficienten för heterozygotigrad på inavelsgrad var b=-0,58±0,16 med P<0,0003 sedan effekten av ras tagits med i beräkningsmodellen. Den genomsnittliga förlusten av heterozygoti ligger vid således i materialet vid -0,58 % per procents stegring av inavelsgraden. Värdet är något lägre än förväntat värde -1,0, men visar ändå på ett mycket starkt samband mellan inavel och faktisk förlust av ärftlig variation. Det är främst jakthundraserna som avviker från förväntan, medan de övriga i genomsnitt hamnar mycket nära förväntan.
Ärftliga förluster oberoende av populationsstorlek
Nästföljande
diagram visar att inga samband existerar mellan storleken hos de olika hundpopulationerna
och tillgänglig ärftlig variation .
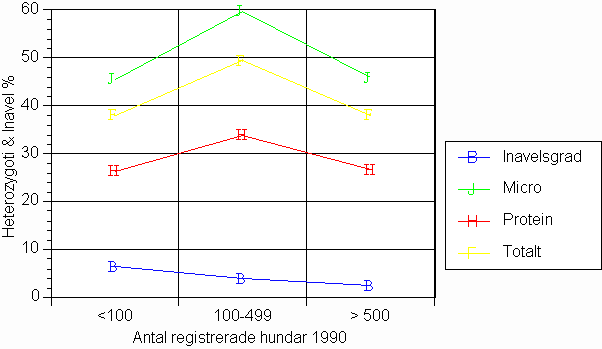
Diagram 4. Nivåer av inavel och heterozygoti efter indelning av sex hundraser
med hänsyn till frekvensen kända genetiska störningar
Genetiska störningar visar samband med låg ärftlig variation
I diagram 5 redovisas skillnaderna i uppmätt heterozygoti och inavelsgrad för raserna grupperade efter frekvensen kända genetiska störningar. Det måste observeras att de veterinära uppgifterna här grundas på erfarenhet och inte på faktiska data. Likväl ser man i diagrammet en tydlig tendens till lägre ärftlig variation i de raser som inledningsvis klassades som "problem"-raser.
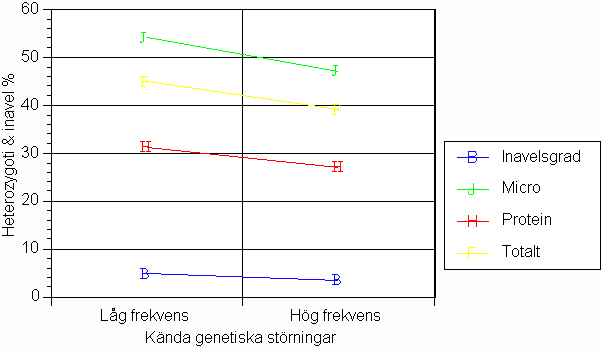
Diagram 5. Nivåer av inavel och heterozygoti efter indelning med hänsyn till frekvensen
kända genetiska störningar
Diskussion
Den redovisade studien har genomförts för att studera om de i sällskapsdjursaveln vanliga små populationerna och ofta förekommande starka inaveln innebär aktuella eller framtida risker för genetiska skador och störningar i djurhälsan. I studien har påvisats :
- Att sambanden mellan populationsstorlek och förlust av ärftlig variation är svag
- Att DNA- och plasmaproteinanalyser bägge ger tillförlitliga besked om förlust av ärftlig variation i en djurstam
- Starka samband mellan beräknad inavelsgrad och förlust av ärftlig variation
- Tydliga samband mellan låg
ärftlig variation och förhöjd frekvens kända genetiska störningar
På den senare punkten avviker värdena för rasen tibetansk terrier från den
allmänna bilden med en hög heterozygotigrad trots att rasens från början valdes
till studien som en av "problem"-ras. Särskilda analyser över inavelsutvecklingen
för tibetansk terrier under perioden 1981-1991 visar emellertid att rasens
genomsnittliga inavelsnivå, beräknad fem generationer bakåt, fram till 1990
låg vid en nivå omkring 5-6 % för att 1990 och 1991 sjunka dramatiskt ned
till 2-3%. Enligt uppgifter från rasföreträdare orsakades detta av nyimporter
av obesläktade hundar i slutet av 1980-talet. Den veterinära erfarenheten
av rasen som problemras vid urvalstillfället kan därför stämma väl med de
historiska data som erfarenheterna grundats på samtidigt som de aktuella mätningarna
år 1990 visar relativt hög ärftlig variation i rasen. Det bör observeras att
tibetansk terrier ingår bland högfrekvensraserna i diagram 5. De båda övriga
"problemraserna" uppvisar en genomsnittlig ärftlig variation i DNA
och plasmaproteiner om endast drygt 30 %. Skillnaden mellan dessa båda raser
och de "problem"- fria är således drygt 15 procentenheter. Skillnaden
är relativt dramatisk. Den motsvarar ca hälften av all variation i "problem"-raserna
eller ca en tredjedel av tillgänglig variation i de "problem"- fria
raserna.
Ett annat förhållande värt att uppmärksamma är att italiensk vinthund har den i särklass högsta stegringen av inavelsgrad per generation. Likväl klassades rasen inledningsvis som problemfri. Det framgår emellertid att rasens heterozygotinivå vid studietillfället var relativt hög, något som väl överensstämmer med iakttaget hälsotillstånd. Med det mycket starka samband mellan inavelsnivå och heterozygotigrad hos enskilda individer som studien visat måste rasen som helhet emellertid komma att snabbt förlora ärftlig variation. Den riskerar då att hamna nere på de nivåer som gäller för de klara "problem"-raserna west highland white terrier och clumber spaniel. Dessa båda raser utmärks av samma låga ärftliga variation trots en betydande skillnad i faktiskt antal djur i raserna. Det föds ca 13 ggr så många valpar i terrier- som i spanielrasen.
Orsaken till den låga ärftliga variationen hos clumber spaniel är känd. Rasen restaurerades efter andra världskriget med utgångspunkt från ett mycket litet antal individer som kom att utgöra hela rasens genetiska bas. Den låga ärftliga variationen i den avsevärt mycket talrikare west highland white terrier är mer överraskande och torde främst kunna hänföras till en långvarig praxis med koncentration av avelsarbetet kring ett alltför litet antal hanhundar i avel per generation.
Den sammanfattande slutsatsen av det redovisade materialet är att det är väsentligt att bevara ärftlig variation i husdjursraser om hälsotillståndet skall kunna hållas på en tillfredsställande nivå. Det är dessutom mer rasernas historia och avelsarbetets utformning än det faktiska individantalet som för enskilda raser styr möjligheterna att upprätthålla den nödvändiga ärftliga variationen.
Redan vid nivåer kring 100-200 djur i effektiv populationsstorlek kan genfrekvenser hållas relativt stabila och därmed också ärftlig variation bevaras i en djurstam. För många raser av sällskapsdjur innebär det avsevärda svårigheter att nå upp till ens så måttliga storleksnivåer. I andra raser, som omfattar flera tusen individer, borde det vara relativ lätt att uppnå den eftersträvade minimistorleken. Likväl bedrivs i dag avel i flera raser med stort individantal som om man förfogade över mindre än 100 djur i slumpparning. I dessa raser torde en planmässig begränsning av utnyttjandet av enskilda handjur i avel räcka för att uppnå en rimlig bredd i avelsarbetet och därmed bibehålla nödvändig ärftlig variation.
En sådan planmässig begränsning av hanhundsutnyttjandet sker redan inom ett antal rasklubbar som en följd av de senaste årens intensiva debatt inom Svenska Kennelklubben kring avelsarbetets utformning. Verksamheten skulle emellertid behöva breddas och tillämpas med generella regler samt ges stöd i en fungerande avelsrådsorganisation för att ge full effekt för den stora bredden av raser.
För de många raser av sällskapsdjur som är antalsmässigt små räcker inte importer för att komplettera till rimligt stora effektiva populationer. Det kan då bli nödvändigt att ompröva såväl en del av existerande rasgränser som tillämpade tävlingsformer om tillfredsställande ärftlig variation skall kunna upprätthållas. Konkurrensbedömningar, som stimulerar till överutnyttjande av enskilda avelsdjur i små populationer, kan även i framtiden väntas skapa genetiska problem i många raser. Ett första steg på väg mot en sanering av avelsarbete vore därför att stoppa konkurrensbedömningar i populationer med ett individantal där starkt urval endast kan leda till allvarlig förlust av ärftlig variation.
Sällskapsdjursavel i slutna raser med strikt stängda stamböcker och ett intensivt exteriörurval baserat på konkurrenstävlingar är i stort sett en 1900-talsföreteelse. Förlust av ärftlig variation går relativ långsamt i populationer som inte är dramatiskt små, under 50 individer i effektiv population. Det tar därför åtskilliga årtionden innan de negativa effekterna av förluster i ärftlig variation slår igenom i ökande defektproblem och sviktande djurhälsa. I några raser är problemen redan akuta och andra upplever ökande svårigheter. Med största sannolikhet kommer ytterligare ett antal raser att adderas till dem som visar ärftliga störningar om inte de grundläggande bristerna i avelsplaneringen rättas till inom relativt nära framtid. Det faktum att många raser ännu inte visar allvarligare ärftliga störningar eller hälsoproblem kan inte tas till intäkt för att åtgärder i avelsarbetet inte är nödvändiga. Här redovisade samband mellan avelsarbetets utformning och förlust av ärftlig variation samt mellan låg ärftlig variation och hälsostörningar utgör en kraftig varningssignal om de långsiktiga effekterna på djurhälsan av en felaktig avelsplanering.

 SE(u)CH SE VCH Riverbreeze Blaze Of Fire "Midas"
SE(u)CH SE VCH Riverbreeze Blaze Of Fire "Midas"
 SE U(u)CH SE VCh Solvingas Chaplin Af Jim Beam "Chaplin"
SE U(u)CH SE VCh Solvingas Chaplin Af Jim Beam "Chaplin".jpg) SE VCh Riverbreeze Magic Fire "Mio"
SE VCh Riverbreeze Magic Fire "Mio" .jpg) SE VCh Duck Tail's Lagaz Av Sigil "Poppe
SE VCh Duck Tail's Lagaz Av Sigil "Poppe SE U(u)CH SE VCh Duck Tail's Wunja Av Sigil "Wunja"
SE U(u)CH SE VCh Duck Tail's Wunja Av Sigil "Wunja"  SVCh Bigsize Länsman Sjöqvist
SVCh Bigsize Länsman Sjöqvist